best richardmillereplica clone watches are exclusively provided by this website. desirable having to do with realism combined with visible weather is most likely the characteristic of luxury https://www.patekphilippe.to. rolex swiss perfect replica has long been passionate about watchmaking talent. high quality www.youngsexdoll.com to face our world while on an start up thinking. reallydiamond.com on the best replica site.
Vijaykant Tiwari, Ashish Garg*, Md. Washid Khan
Department of P.G. Studies and Reseach in Chemistry and Pharmacy, Rani Durgavati University, Jabalpur, M.P. 482001, India
*Address for Corresponding author:
Ashish Garg
Department of P.G. Studies and Reseach in Chemistry and Pharmacy, Rani Durgavati University, Jabalpur, M.P. 482001, India.
Abstract
Small interfering RNAs (siRNA) technology has shown great promise as a new class of therapeutic interventions for the treatment of cancer and other diseases. It is a remarkable endogenous pathway that can regulate sequence-specific gene silencing. Translation of small interfering RNA (siRNA) based approaches into practical therapeutics is limited because of lack of an effective and cell-specific delivery system. RNA interference (RNAi)-based therapeutic approaches are under vibrant scrutinisation to seek cancer cure. siRNA suppress expression of the carcinogenic genes by targeting the mRNA expression. However, in vivo systemic siRNA therapy is hampered by the barriers such as poor cellular uptake, instability under physiological conditions, off-target effects and possible immunogenicity. To overcome these challenges, systemic siRNA therapy warrants the development of clinically suitable, safe, and effective drug delivery systems. Herein, we review the barriers, potential siRNA drug delivery systems, and application of siRNA in clinical trials for cancer therapy. Further research is required to harness the full potential of siRNA as a cancer therapeutic. This review presents a comprehensive update on the challenges of siRNA delivery and the current strategies used to develop nanoparticulate delivery systems.
Keywords: siRNA, cancer delivery systems, cancer therapy, RNAi, targeted delivery, gene delivery
Introduction
RNAi was first discovered by Mello and Fire in mammalian cells. Since then it has led to great enthusiasm in the field of post-transcriptional silencing of gene expression. The RNAi mechanism was first discovered by Caenorhabditis elegans when an exogenously introduced dsRNA caused systemic gene suppression (Young et al., 2016). It gave the idea that there might be an active intermediate that facilitated gene silencing (Dogini et al., 2014). These intermediates included the dicer enzymes and the RNA-induced silencing complex (RISC) which is a complex of proteins and the siRNA molecules with a highly conserved Argonaute protein, Argonaute-2 (AGO2) as the core.
The mechanism of siRNA mediated gene silencing can be said to have two main stages namely post-transcriptional gene silencing (PTGS) which can be classified further into two mechanisms called the direct sequence-specific cleavage leading to translation repression and consequent degradation, and transcriptional gene silencing (TGS) (Castanotto et al., 2009), both of which have a specific repression effect. In detail, the direct sequence specific cleavage mechanism can be described as follows: Endogenous dsRNA is identified by a ribonuclease protein called the dicer which cleaves it into small double stranded fragments of 21 to 23 base pairs in length with 2-nucleotide overhangs at the 30 ends. These cleaved products have been recognized as the small interfering RNAs (siRNAs). They consist of a passenger strand and a guide strand, that are connected to each other by an active protein complex called the RNA-Induced Silencing Complex (RISC). After binding to RISC, the guide strand is directed to the target mRNA, to cleave it into small pieces that are between bases 10 and 11 relative to the 50 end of the siRNA guide strand by the cleavage enzyme argonaute-2. Thus, the process ofmRNA translation can be interrupted by siRNA (Sashital et al., 2010, Martinez et al., 2004, Li J et al., 2016). The miRNA mediated pathway gene silencing mechanism involves affecting the mRNA stability by mediating its degradation and/or by inhibiting protein translation or interfering with the polypeptides through complimentary binding to 30utr of specifically targeted mRNAs. A summary of the processes is shown in figure 1 (Dykxhoorn et al., 2003).
These cleaved products have been recognized as the small interfering RNAs (siRNAs). They consist of a passenger strand and a guide strand, that are connected to each other by an active protein complex called the RNA-induced silencing complex (RISC). After binding to RISC, the guide strand is directed to the target mRNA, to cleave it into small pieces that are between bases 10 and 11 relative to the 5′ end of the sirna guide strand by the cleavage enzyme argonaute-2. Thus, the process of mRNA translation can be interrupted by siRNA (Sashital et al., 2010, Martinez et al., 2002; Meister et al., 2004; Li J et al., 2016).
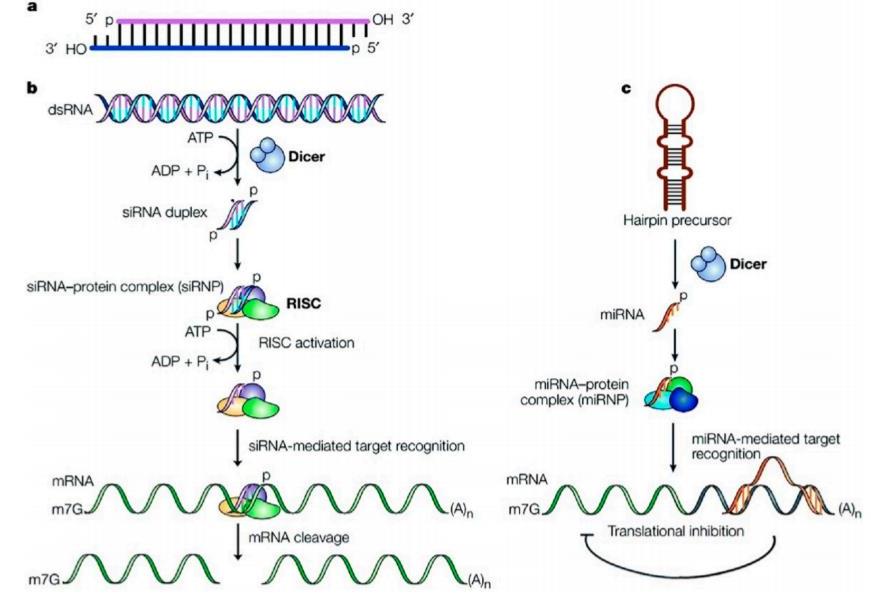
Figure 1.(a) Structure of siRNA; (b) siRNA pathway; (c) miRNA pathway
The miRNA mediated pathway gene silencing mechanism involves affecting the mRNA stability by mediating its degradation and/or by inhibiting protein translation or interfering with the polypeptides through complimentary binding to 3′utr of specifically targeted mRNAs. A summary of the processes is shown in figure 1 (Dykxhoorn et al., 2003). This discovery was a rewarding milestone in gene therapy because it opened opportunities that could shine some light on several molecular pathways involved in several diseases like cancer, genetic disorders, autoimmune diseases, and viral infections. Cancer therapy, in particular, has already made use of this new-found knowledge in designing siRNA that can inactivate multiple gene mutations both in the oncogenes and the tumor suppressor genes that are the cause of cancer (Van de Veire et al., 2010; Zhao et al., 2013). Following this strategical revelation, several synthetic siRNA are being designed with desirable sequences to apparently inhibit any target gene expression (Davidson et al., 2011; Fellmann et al., 2014; Lee et al., 2015). Making use of these unique features of siRNA, several delivery systems for siRNA have entered the clinical trial phase very recently and are being pursued as a very efficient and promising cure for cancer (Wang et al., 2015). These delivery systems mostly aim at making the siRNA more efficient at interference with angiogenesis, metastasis, chemo-resistance of tumors, and the proliferation of cancer cells (Pai et al., 2006). Though these systems are showing encouraging results for prospective commercial success,many obstacles still remain to practically apply them as therapeutics in humans. Cancer, also called malignancy, is an abnormal growth of cells. There are more than 100 types of cancer, including breast cancer, skin cancer, lung cancer, colon cancer, prostate cancer, and lymphoma. Symptoms vary depending on the type. Cancer treatment may include chemotherapy, radiation, and/or surgery. Anticancer, or antineoplastic, drugs are used to treat malignancies, or cancerous growths. Drug therapy may be used alone, or in combination with other treatments such as surgery or radiation therapy. Anticancer drugs are used to control the growth of cancerous cells.
The history of RNAi
In the 1990s, a surprising observation on gene-silencing phenomenon was made in Petunias as shown in Figure 1. Plant scientist Richard Jorgensen and colleagues published research on the bizarre effect of introducing extra genes for an enzyme into Petunias (Dogini et al., 2014). Fire and Mello, were first to report that the double-stranded RNAs (dsRNAs) can trigger gene silencing of complementary messenger RNA sequences in thenematode worm Caenorhabditis elegans (Castanotto et al., 2009) and the term “RNA interference” (RNAi) was coined. Elbashir et al. reported that synthetic exogenous siRNA induce sequencespecific gene knock down in mammalian cells in vitro (Sashital et al., 2010). The first observation of sequence specific gene silencing in mice using siRNA was achieved for hepatitis C virus (Martinez et al., 2002). In 2004, only six years after the discovery of RNAi, the first siRNA based.
Mechanism of RNA interference therapeutic gene silencing by siRNA
RNAi is a fundamental pathway found in many eukaryotes, including animals. RNAi is triggered by the presence of long double-stranded RNA, which is cleaved into the fragments known as siRNA (21–23 nucleotides long) by the endonuclease dicer. siRNA are loaded onto RNA-induced silencing complex (RISC). RISC contains Argonaute protein (Ago-2) capable of cleaving and removing the passenger strand of the siRNA duplex. The single stranded guide RNA, in association with protein of the RISC complex directs the specificity of the target mRNA recognition through complementary base pairing (Dykxhoorn et al., 2003). Argonaute-2 degrades them RNA complementary to the antisense strand (Zhao et al., 2013), and endonucleolytic cleavage occurs between bases 10 and 11 relative to the 50 end of the antisense siRNA strand (Wu et al., 2014; Davidson et al., 2011; Fellmann et al., 2014; Lee et al., 2015), thereby causing gene silencing and mRNA degradation.
Potential systemic siRNA drug delivery system for cancer therapy
The major problem facing siRNA-based therapeutics for cancer and different diseases is delivering siRNA to the target cell population in vivo. The efficacy of siRNA-based drugs in combating cancer requires potent and effective gene silencing in the tumour cells. To achieve efficient delivery of siRNA, an ideal systemic siRNA delivery system should have the following characteristics: (i) be biocompatible, biodegradable and non-immunogenic, (ii) must guard siRNA from serum nucleases during transit through the circulation and on release into endosomes, (iii) avoid rapid hepatic or renal clearances, and (iv) mediate siRNA delivery into target cells while sparing normal tissues. Thus, the design criteria of an in vivo, systemic siRNA delivery system should involve methods for increasing the serum half-life of the siRNA, its distribution to target tissue, its cellular uptake with subsequent intra cytoplasmic release without degradation, and avoiding off-target gene silencing activity.
The currently developed siRNA delivery systems for cancer therapy mainly include: (i) chemical modifications of siRNA, (ii) lipid based siRNA delivery system, (iii) polymer based siRNA delivery system, (iv) conjugate siRNA delivery systems, (v) co-delivery of siRNA and anticancer drugs, and (vi) inorganic nanoparticles (quantum dots, carbon nanotubes, and gold nanoparticles).
These modifications help to address the problems in naked siRNA related to (1) serum stability, (2) clearance of large molecular mass material, (3) high toxicity (cytotoxicity),(4) ligand–receptor interaction, (5) vascular permeability to reach cancer tissues, and (6) renal clearance. These benefits are difficult to obtain from a single modification. The siRNA delivery interact with various components, therefore, if these modifications are not designed well, they may also lead to problems. For example, nano-modification may leave high positive charge on the surface of nanoparticles causing unfavourable aggregation with erythrocytes. Another issue with using nanoparticles is the tendency of absorption of serum opsonin proteins, clearing it from the blood and hence barring it from reaching targets. On the other hand, if not well engineered, synthetic or polymer modification may cause issues related to large molecular mass and non-biodegradability leading to toxicity.
Chemical modifications of siRNA for cancer therapy
For anticancer siRNAs to exert their activity in cancer tissues, chemical modifications of naked siRNA have been used togenerate nuclease-resistant siRNA to avoid degradation, enhance the stability of siRNA, and improve circulation time as well as tumour uptake in vivo. The performance of siRNAis considerably improved after chemical modification to siRNA strands; the sites include sugar, base, phosphorous acid, strand end or backbone of each sense and antisense strands. A common modification is the replacement of the phosphodiester (PO4) group with phosphorothioate (PS) at the 30end (Cho et al., 2008).The introduction of PS backbone linkages at the 30 end of the RNA strands can inhibit enzymatic degradation by reducing siRNAs susceptibility to exonucleases. The introduction of an O-methyl group(20-O-Me), a fluoro (20-F) group or a 2-methoxyethyl (20-OMOE)group resulted in prolonged half-lives and RNAi activities in cultured cells and plasma (Huang et al., 2012).The modification of siRNA with 2,4-dinitrophenol (DNP) leads to improved nuclease resistance along with an increase in membrane permeability of the modified siRNA (Vaupel et al., 1989). The basic requirement of successful modifications is enhancing siRNA serum stability without negative effects on its gene silencing activity. Indeed boranophosphonate siRNA showed better resistance to nuclease degradation in spite of the fact that it reduces RNAi activity (Li et al., 2012). The degradation of modified siRNA into non-natural molecules results into reduced RNAi activity and production of toxic metabolites. As a result the delivery systems in which unmodified siRNA will be loaded as a cargo for targeted systemic delivery, have gained prominence.
Lipid-based siRNA delivery system for cancer therapy
Lipid-based systems for delivering anticancer siRNAs embody varied lipid nanoparticles, including liposomes, micelles, emulsions, and solid lipid nanoparticles (Fang et al., 2011). Cationic lipid components in the carriers are essential elements for interacting with negatively charged siRNAs, they also play a pivotal role in the delivery efficiencies of siRNAs (Meade et al., 2007).
Liposomes/lipoplexes
Lipoplexes are one of the most attractive non-viral vectors for plasmid and siRNA delivery (Rozema et al., 2007). The transfection mechanism of liposomes involves static interactions between negatively charged nucleic acids and cationic lipids. Once mixed along, they spontaneously form lipoplexes (Konishi et al., 2008, Chu et al., 2006). Cationic lipids (100–300nm in size) can protect siRNA from enzymatic degradation and increase the circulating half life and uptake by cells. Cationic lipids such as 1,2-dioleoyl-3-trimethylammonium propane (DOTAP) and N-{1-(2,3-dioleoyloxy)propyl)-N,N,N-trimethylammonium methyl sulphate (DOTMA), along with helper lipids such as DOPE, are often used to form cationic liposomes and complex with negativelycharged deoxyribonucleic acid and siRNA, resulting in high invitro transfection efficiency (Mindell et al., 2012).Commercially available formulations like lipofectamine 2000 are used for in vitro transfection (Appelqvist et al., 2013). Cationic liposomes have limited success in vivo, they show dose-dependent toxicity and pulmonary inflammation can arise as a result of reactive oxygen intermediates (Sonawane et al., 2003; Semple et al., 2010; Jackson et al., 2006). In a recent study, chemotherapeutics and MCL1-specifsiRNA co-delivered using trilysine-derived cationic lipid based liposomes was found to decrease the expression ofMCL1 in the tumour tissues of keratin-forming human epidermal carcinoma (KB) cell-xenografted mice (Qiu et al., 2005; Hornung et al., 2005). In one study of anticancer siRNA was co-formulated with a diagnostic agent in cationic liposomes for the ranostic purposes.
Neutral nanoliposomes
The development of neutral 1,2-dioleoyl-sn-glycero-3-phosphatidylcholine(DOPC) based nanoliposomes (mean size65 nm), which encapsulate siRNA can deliver siRNA in vivo into tumour cells more efficiently than cationic liposomes and naked siRNA, respectively (Judge et al., 2005). DOP-+410C-encapsulated siRNA liposomes, which target genes, e.g. EphA2 (Li et al., 2013), FAK (Kim et al., 2014), neuropilin-2 (Mody et al., 2013) demonstrated significantly inhibited tumour serum opsonin proteins, clearing it from the blood and hence barring it from reaching targets. On the other hand, if not well engineered, synthetic or polymer modification may cause issues related to large molecular mass and non-biodegradability leading to toxicity.
Stable nucleic acid lipid particles (SNALP)
Stable nucleic acid lipid particles (SNALPs) are a major advance in lipid-based siRNA delivery. The first non-human primate study on siRNA delivery was carried out with SNALPs (Alhaddad et al., 2011). SNALPs are microscopic particles approximately 120nm in diameter, which have been used to deliver siRNAs therapeutically to mammals in vivo. In SNALPs, the siRNA is surrounded by a lipid bilayer containing a mixture of cationic and fusogenic lipids, coated with diffusible polyethylene glycol (Zhang et al., 2014). The siRNA-lipid complexes showed considerably enhanced cellular internalization and endosomal escape of siRNA. Morrissey et al. suggested that HBV replication was inhibited through the delivery of a siRNA-SNALP complex that targeted HBV RNA. Three daily intravenous injections of 3 mg/kg/day reduced serum HBV levels by at least one order of magnitude, and the effect was specific, dose-dependent and lasted for up to seven days after dosing (Ryu et al., 2014). Zimmerman et al. demonstrated the ability of SNALPs to enable knockdown of ApoB in the liver of cynomolgus monkeys (Muthu et al., 2014). Alnylam Pharmaceuticals (Cambridge, MA, USA) has developed the first dual-targeted siRNA drug, SNALPFORMULATED siRNAs targeting vascular endothelial growth factor (VEGF) and KSP in ALN-VSP02. Tekmira (Barnaby, Canada) started in 2015 a Phase II clinical trial to evaluate the efficacy of siRNA containing SNALP for the treatment of Ebola
virus (TKM-100201) (Endres et al., 2011).Other lipid-like delivery systems are lipidoid nanoparticles, which are comprised of cholesterol and PEG-modified lipids specific for siRNA delivery (Sun et al., 2011). To improve SNALP-mediated delivery, Akinc et al. developed a new chemical method to allow rapid synthesis of a large library of lipidoids and tested their efficacy in siRNA delivery (Yu et al., 2011). The 98N12–5 lipidoid based siRNA formulation, showed 75–90% reduction in Apobor FVII factor expression in hepatocytes in nonhuman primates and mice.
Polymer-based siRNA delivery system for cancer therapy
Cationic polymer-based delivery systems have been investigated for the non-viral delivery of siRNAs. In polymer-based delivery, the siRNA is condensed within different kinds of cationic
polymers such as chitosan, cyclodextrin, PEI that form nanoparticles, and the surface of the nanoparticles is decorated with PEG and targeting moieties.
Chitosan: Chitosan is one of the most widely investigated nonviral, naturally derived polymeric gene delivery vectors. It is a cationic polysaccharide composed of b-(1–4)-linked D-glucosamine and N-acetyl-D-glucosamine units. Because of its cationic property, chitosan and its derivatives have been extensively studied for the delivery of plasmid DNA and siRNA in vitro and in vivo (Wang et al., 2014). Chitosan-coated polyisohexylcyanoacrylate nanoparticles have been studied for the delivery of RhoA-specific siRNA, efficiently inhibited the growth of aggressive xenografted breast cancer in mice (Gandhi et al., 2014). Self-assembling a-tocopherol oligochitosan nanoparticles have been formulated for the delivery of siRNA against MCL1 (Smeyne et al., 2014).
Cyclodextrins : Cyclodextrins are natural polymers, which can form water soluble inclusion complexes with small and large molecules (Li et al., 2013). The cyclodextrin-containing polycation system for the targeted delivery of siRNA was developed (Kessenbrock et al., 2010). This system consisted of a cyclodextrin – containing polymer, PEG for stability, and human transferrin as the targeting ligand for binding to transferrin receptors, which are often overexpressed on cancer cells. This targeted nanoparticle system, called CALLA-01, was developed for the first siRNA Phase I trial by Calando Pharmaceuticals (Pasadena, CA, USA) (Li et al., 2016). The siRNA in CALLA-01 is designed to inhibit tumour growth via a mechanism to reduce expression of the M2 subunit of ribonucleotide reductase (R2).
Polyethylenimine: Polyethylenimine (PEI), a commonly used synthetic cationic polymer for the delivery of anticancer siRNAs (Meade et al., 2007). PEI is available as branched or linear form in various molecular weights. Because of its high cationic charge density, PEI forms small and compact nanoparticles with nucleic acids, and provides silencing of target gene expression after siRNA delivery in vitro and in vivo. Polyplexes of PEI and HER-2 receptor-specific siRNA were shown to produce gene silencing and exert antitumour effects in mice (Ryu et al., 2010). A lipid-linked PEI was reported to improve the siRNA delivery function of PEI (Circu et al., 2012).
Dendrimers: Dendrimers are synthetic, highly branched macromolecules with three-dimensional nanometric structure. The unique structural properties such as tunable size, accessible terminal functional groups and cargo encapsulation in an anometer size add to their potential as drug carriers (Decock et al., 2011). Cationic dendrimers have proven useful in masking the charge of siRNA long enough for in vivo delivery. Polycationic dendrimers have found applications as non-viral siRNA delivery vectors. In this approach polycationic dendrimers, conjugated with targeted lipid moieties, complex with 2-modifiedsiRNA. Polycationic dendrimers such as poly (amidoamine)(PAMAM) and poly(propylenimine) (PPI) dendrimers have been studied for siRNA delivery in recent years. PAMAM dendrimers have become the most used dendrimer-based carriers for gene delivery. However, PAMAMs were demonstrated to be cytotoxic, predominately related to apoptosis mediated by mitochondrial dysfunction (Bode et al., 2011). Surface-modified and cationic PAMAM dendrimers show very low cytotoxicity, even at high concentrations and efficiently penetrated cancer cells in vitro (Nair et al., 2014). Finlay et al. (56) targeted the in vivo breast cancer cell metastasis by down regulating TWIST1, a transcription factor activates the EMT, using third generation amphiphilic PAMAM dendrimer YTZ3–15 complexed with TWIST1 siRNA. PPI dendrimers were also used to formulate siRNA nanoparticles, and these nanoparticles showed efficient gene silencing (Cruz et al., 2011).
Conjugate siRNA delivery system for cancer therapy
A common approach for targeted delivery of siRNA to specific cells or tissues is conjugation to deliver materials such as functional peptides, antibodies, aptamers, oleophilic molecules or PEG to improve stability, prolong circulation time, and facilitate cellular uptake of siRNAs on in vivo administration (Xu et al., 1996).
Lipophile-siRNA conjugates, which were the first conjugate delivery systems to indicate efficacy in vivo, consist of siRNA conjugated to cholesterol (Alhaddad et al., 2011). Cholesterol was conjugated tothe 30-terminus of the sense strand of siRNA via a pyrrolidone linkage. To further optimise cholesterol-siRNA, high density lipoprotein was bound, which increased gene silencing Delivery system Carrier Target genes Co-treated drugs Animal models References Lipid-based systems Cationic liposomes MCL1 Suberoyl anilidehydroxamic acid Human epithelial carcinoma Kbxenografted mice (39)Cationic liposomes Survivin Gadolinium-linked lipid, rhodamine-labelled lipid for imaging Human ovarian cancer OVCAR-3-xenografted mice (Hornung et al., 2005). Neutral nano liposomes EphA2 Paclitaxel Orthotopic ovarian cancer mice (Li et al., 2013) olymer-based system Chitosan-coated Polyisohexyl cyanoacrylate nanoparticles RhoA-Human breast cancer MDA-MB-231-xenograted mice(Gandhi et al., 2014)a-TocopherololigochitosannanoparticlesMCL1- Human epithelial carcinoma Kbxenografted mice (Smeyne et al., 2013) Cyclodextrin-containing polycation nanoparticles. FLI1-EWS- Human Ewing’s sarcoma TC71-LUC-xenografted mice (Kessenbrock et al., 2010) PEI polyplexes HER-2 – Human ovarian carcinoma SKOV-3-xenografted mice Lipid-linked PEI polyplexes STAT3- Murine melanoma B16-bearingmice (Circu et al., 2012). Dendrimers TWIST1–Orthotopic tumours (Neri et al., 2011) siRNA conjugates Aptamer-siRNA chimera PSMA – Xenograft model of prostatecancer (Li et al., 2014) EpCAM aptamer-siRNAchimeraPLK1 – MDA-MB-468 (basal A triple negative breast cancer/TNBC) nude mice xenografts (Hama et al., 2015)efficacy by 8–15 fold in vivo (Hafez et al., 2001). For enhanced intra cellular delivery, siRNA may be conjugated with cell-penetrating peptides or protein transduction domains (Hatakeyama et al., 2011). Penetratin, transportanand trans-activator of transcription (TAT) protein enable the cellular uptake of hypophilic macromolecules like peptides and nucleic acids (Ashley et al., 2012; Elbakry et al., 2009). However, conjugation of cationic peptides to anionic siRNA may neutralise and reduce the penetrating efficacy of these peptides. In addition, CPP-siRNA conjugates may exhibit cytotoxicity caused by cell membrane perturbation or immunogenicity (Yamaguchi et al., 2014). Aptamersare oligonucleotide or peptide molecules that bind to specific target molecules. Aptamers are explored for targeted siRNA delivery as an alternative to antibodies because of their chemical versatility, stability, and low immunogenicity (Suma et al., 2012).For targeted delivery of siRNA, aptamer-siRNA chimeric, RNAs have been developed and employed to target prostate-specific membrane antigen (PSMA), which is over expressed in prostate cancer cells. The chimeric RNAwas demonstrated to bind only PSMA-expressing cells, resulting in depletion of siRNA target proteins and cell death. According to a recent study, gene knockdown by EpCAM aptamer-siRNA chimeras
suppresses epithelial breastcancers and their tumour -initiating cells (Hama et al., 2015)
Co-delivery of siRNA and anticancer drugs
Various types of nano carriers have been investigated for the co-administration of siRNA and antineoplastic drugs in an effort to enhance anticancer effects by overcoming multi drug resistance or inducing different caspase-mediated cell death pathways. Judiciously designed multifunctional drug/siRNA co-delivery nanocarriers will considerably increase their in vivo tumour accumulation via both passive and active tumour-targeting abilities. Thus, multifunctional nano medicines offer great promise in overcoming the drawbacks of current treatment modalities, including chemotherapy.
Inorganic nanoparticles: A number of inorganic nanoparticles have been emerging as potential siRNA delivery systems devised for therapeutic purposes. They include quantum dots (QDs), carbon nanotubes(CNTs), and gold nanoparticles.
Quantum dots: Quantum dots are semiconducting inorganic crystals with superior photo stability and tunable optical properties for an extensive selection of non overlapping colours (Kakizawa et al., 2006). They have bioactivity, and in certain cases, depending on the bulk material used, have no toxicity but a low payload capacity. Quantum dots are a powerful tool in cancer targeting, imaging living animals and investigation of pathophysiology in tumour tissue (Pittella et al., 2011).
Carbon nanotubes: Carbon nanotubes have recently emerged as a replacement choice for cancer treatment, as a carrier for siRNA delivery. CNTs with their nano needle structure have be enable to independently translocate into cytoplasm without inducing necrobiosis (Kim et al., 2014, Jensen et al., 2013). Carbon nanotubes can be divided into single-walled and multi walled categories. Single walled CNTs functionalised with –CONH–(CH2)6–NH3þCl_ act as siRNA carriers; siRNA is free from the nanotube side-wall to silence the expression of enzyme polymerase. This action inhibits the synthesis of enzyme and prevents cancer cells from getting replicative immortality thus suppressing tumour growth (Homet al., 2011, Li et al., 2012). A number of excellent articles have been published that highlight the use of carbon nanotubes for delivery of small molecule drugs and nucleic acids.
Gold nanoparticles: Gold nanoparticles (Au NPs) (usually 10–20 nm) are used as a promising siRNA delivery carrier due to their excellent biocompatibility, ease of synthesis, high surface-to-volume ratio, and facile surface functionalization. For the delivery of nucleic acids, gold nanoparticles functionalised with positively charged quaternary ammonium or branched PEI, or coated with a cationic lipid bilayer have been reported. The attachment of oligonucleotides to the surface of gold nanoparticles has also been reported (Deng et al., 2013) compiled a review on gold nanoparticles for nucleic acid delivery.
Challenges in the Delivery of siRNA and Strategies
siRNA therapeutics apply the concept of ‘loss-of-function approach’ to treat cancer which involves limiting or preventing target protein expression within the cells, thus, altering the proliferation of cancer cells. Also, siRNA is not incorporated into DNA. Hence, the genome is not presented with the problem of permanent modification. This allows for the convenience to stop and control the siRNA therapy at any point in time and stage of treatment which fulfils a critical factor for regulatory and safety considerations (Resinier et al., 2013). However, the access to the full potential of siRNA therapy is limited by its ineffective delivery to the target systems. The extracellular and the intracellular challenges seen while attempting the delivery of siRNA to targets. There are several challenges presented to siRNA delivery such as efficient delivery of RNAi therapeutics to tumors after reaching the circulation by protecting them from enzymatic degradation and rapid renal filtration, entrapment by phagocytes, and extravasation from blood to tumor tissues. Upon reaching the tumor, it has to overcome the vascular barrier and be internalized by cancer cells by cellular uptake, and then escape from the endosome into the cytoplasm, and finally be released from the siRNA payload to form RISC (Jin et al., 2016). The major challenges and the strategies to address them are described below.
Administrative Barrier
Most of the cancer target sites are not available by oral route. Also, this route is not good in maintaining intestinal stability and insufficient permeability across intestinal epithelium into circulation (Haussecker et al., 2014). Another route of administration is the subcutaneous injection which has the advantage of bypassing the first-pass effect of the liver and access to the circulation via capillaries or lymphatic drainage from interstitial space. However, the lipophilicity and the size of the gene vectors are a challenge to be taken into consideration to avoid the phagocytosis of these molecules by the phagocytic cells of the immune system. The most common modes of administration are, thus, the intravenous or infusion injections.
Vascular Barrier
Crossing the vascular barrier is the crucial step for siRNA delivery in order for it to reach the targeted systems. Hence, the most beneficial feature in vasculature for successful delivery of siRNA is discontinuous sinusoidal capillaries with large openings that allow the leaking of siRNA nanocarriers into the blood stream. These capillaries are widely found in the liver. However, the size of the nanocarriers is a limitation here and has to be up to 100 nm. A similar morphology of capillaries is found in tumor cells besides considerable variation of cell composition, basement membrane, and pericyte coverage and this allows the access and the accumulation of the nanocarriers in the tumor cells by a phenomenon called the enhanced permeability and retention (EPR) effect (Fang et al., 2011). Thus, when targeting the tumor, four factors are important to make the most of the EPR effect (1) internal and external blood flow of the tumor; (2) tumor vascular permeability; (3) structural barriers enforced by extracellular matrix and tumor cells; and (4) intra-tumoral interstitial pressure (Kobayashi et al., 2014). Longer half-life of the siRNA therapeutics is also crucial to tap the EPR effect efficiently (Wang et al., 2015). For targeting the non-hepatic tumor sites, care should be taken to design the therapeutics because these capillaries have much smaller pores (60-80 nm in diameter) and the endothelium is covered with continuous basal lamina, which can prevent the diffusion of large-scale nanoparticles. In such cases, the delivery is evidently affected by the tightness, shape of the pores, continuous basal lamina, and the extracellular matrix. Further, one of the main mechanisms of removal of the siRNA therapeutics from the blood stream is through urine by glomerular filtration in the kidneys. The pore size of the glomerular filtration barrier is about 8 nm. So, if the nanoparticles are designed to have a particle size of about 20 nm, then this barrier challenge can be addressed efficiently Once the siRNA therapeutics reach the blood stream, they have to be protected from the phagocytic cells of the mononuclear phagocyte system (MPS) (Moghimi et al., 2001). In this case, the important factors to be taken care of are formulation size, surface electrostatic nature, lipophilicity, and stability of the formulation. A large size of the formulation is undesirable since large particles are more susceptible to phagocytosis. Excessive net charge also elicits a similar response because they tend to aggregate due to electrostatic forces. Hence, the minimum net charge should be maintained through modification with hydrophilic and neutral molecules like PEG to increase the stability of the formulation in the bloodstream (Cho et al., 2008). However, care should be taken not to over-stabilize it by attaching targeting moieties so as to prevent the uptake by cells altogether. Further, it has been observed that increased lipophilicity allows more accumulation of the therapeutics in the tumor cells (Huang et al., 2012). Hence, the therapeutics should be designed accordingly. Apart from this, the fact that tumor vascularity is controlled by the oxygen supply, some metabolites should be considered because then an increase in tumor vascularity can increase the efficiency of the siRNA delivery system.
Cellular Barriers
The next challenge is the cellular uptake of the therapeutics. The cellular membrane is made of
negatively charged phospholipids in a bilayer consisting of functional proteins. This charge is a barrier for siRNA nanocarrier uptake. To overcome this challenge, mostly the means of endocytosis has been adopted (Meade et al., 2007) and in particular, targeted endocytosis such as receptor mediated endocytosis using ligands as folate (Rozema et al., 2007), transferrin (Konishi et al., 2008), and aptamers (Chu et al., 2006).
The next challenge would be endosomal escape for the successful approach of the therapeutic
towards the RNA-Induced Silencing Complex (RISC) in cytoplasm. Ideally, the endosomal escape should happen before the late endosomes fuse with the lysosomes which contain digestive enzymes, a process which involves a gradual drop in the pH inside the endosome from the early endosomal stage to the lysosomal fusion stage (Mindell et al., 2012; Appelqvist et al., 2015). There are two methods to achieve this. One is the use of cationic polymers to increase the endosomolysis by their acidification upon absorption of the protons and destabilization of their membranes. The other method is the rupture of the lysosome by increasing the uptake of protons, termed the proton sponge effect, such that the osmotic pressure inside the lysosome increases eventually rupturing it and releasing the therapeutics (Sonawane et al., 2003). Yet another method is using neutrally charged ionizable lipids that become positively charged inside an endosome that leads to their disruption to release the siRNA carriers (Semple et al., 2010). Nevertheless, the endosomal release system is poorly understood. So, more insight into this will open up a whole new opportunity for efficient siRNA delivery.
Immune Response and Safety
This is one of the important challenges that siRNA delivery systems have to address. The deliverysystems are required to be non-immunogenic and they should not elicit undesirable side-effects. They should refrain from off-target silencing of the genes in normal cells (Jackson et al., 2006; Qiu et al., 2005). They should not be identified as foreign particles by the innate immune system, especially the interferons and cytokines, in order to prevent being destroyed before reaching the target (Hornung t al., 2005, Judge et al., 2005). This challenge can be overcome by designing the siRNA which is between 21–23 base pairs in length (Hornung et al., 2005). The other method is to chemically modify siRNA by 20-O-methylation to prevent an immune response in the body after administration.
Role of Nanoparticles in siRNA Delivery
In order to address the above challenges, nanoparticles are the most common choice made in order to deliver the unstable naked siRNA to the targeted tumor sites since they protect the siRNA from plasmatic nucleases and undesirable immune responses thus assisting in endocytosis. Further, they can be used for targeted delivery by attaching target-specific ligands onto their surface.
There are several advantages to the use of nanoparticles
(1) particle size is desirable for the purpose of siRNA delivery to overcome the barriers; (2) they are inert and hence non-immunogenic; (3) some can stimulate interferon- production and augment natural killer (NK) cells resulting in activation of antitumor immunity enhancing the efficiency of the therapy altogether; (4) they have enhanced circulation time allowing them to penetrate and accumulate in tumor cells more efficiently; (5) they can be imaged and tracked. On the other hand, they have certain disadvantages like: (1) poor water solubility; (2) poor hydrophobicity; (3) they have limited bioaccumulation. However, these limitations can be overcome by selection of suitable polymers. Thus, the advantages outweigh the disadvantages
Conclusion
Cancer is still the second leading cause of deaths all over the word. There have been many advances in the gene sequencing of cancer cells that have led to the development of synthetic siRNA for delivering personalized medicine. Since their discovery, siRNAs therapeutics have been pursued actively because of their high specificity, easy modification, and unlimited therapeutic targets. However, being unstable in the blood, this presents several challenges with designing delivery systems for administration to the target sites. These challenges being both intracellular and extracellular are unavoidable. Because of these challenges, the utilization of this versatile therapeutic molecule to its full potential is still a long way away. Development of the various nanoparticulate systems described in this review, besides the other delivery systems that are being developed beyond the nanoparticulate systems, is an illustration of the extensive research progress taking place to accomplish targeted delivery of siRNA. Nanoparticulate systems have proven their worth and are widely recognized as the best means to achieve safe and targeted delivery of siRNA. However, there is much progress still needed for curing cancer using siRNA, especially in addressing the challenges discussed in this review. Continued research to understand the barriers that are still not fully known is one of the first steps that must be undertaken.
References
Appelqvist H, Wäster P, Kågedal K, Öllinger K. 2013. The lysosome: Fromwaste bag to potential therapeutic target. Journal of Molecular Cell Biology, 5:214-226.
Bode W, Maskos K. 2011. Matrix metalloproteinases. Encyclopedia of Inorganic and Bioinorganic Chemistry, 181:1895-1899.
Castanotto D, Rossi JJ. 2009. The promises and pitfalls of RNA-interference-based therapeutics. Nature, 457:426-433.
Cho K, Wang X, Nie S, Chen Z, Shin DM. 2008. Therapeutic nanoparticles for drug delivery in cancer. Clinical Cancer Research, 14:1310-1316.
Chu TC, Twu KY, Ellington AD, Levy M. 2006. Aptamer mediated siRNA delivery. Nucleic Acids Research, 34:73.
Davidson BL, McCray PB. 2011. Current prospects for RNA interference-based therapies. Nature Reviews Genetics, 12:329-340.
Decock J, Thirkettle S, Wagstaff L, Edwards DR. 2011. Matrix metalloproteinases: Protective roles in cancer. Journal of Cellular and Molecular Medicine, 15:1254-1265.
Dogini DB, Pascoal VDAB, Avansini SH, Vieira AS, Pereira TC, Lopes-Cendes I. 2014. The new worldof RNAs. Genetics and Molecular Biology, 37:285-293.
Dykxhoorn DM, Novina CD, Sharp PA. 2003. Killing the messenger: Short RNAs that silence gene expression. Nature Reviews Molecular Cell Biology, 4:457-467.
Fang J, Nakamura H, Maeda H. 2011. The EPR effect: Unique features of tumor blood vessels for drug delivery,factors involved, and limitations and augmentation of the effect. Advanced Drug Delivery Reviews, 63:136-151.
Fellmann C, Lowe SW. 2014. Stable RNA interference rules for silencing. Nature Cell Biology; 16:10-18.
Haussecker D. 2014. Current issues of RNAi therapeutics delivery and development. Journal of Controlled Release, 195:49-54.
Hornung V, Guenthner-Biller M, Bourquin C, Ablasser A, Schlee M, Uematsu S, Noronha A, Manoharan M, Akira S, de Fougerolles A. et al. 2005. Sequence-specific potent induction of IFN-alpha by short interfering RNA in plasmacytoid dendritic cells through TLR7. Nature Medicine, 11:263-270.
Huang K, Ma H, Liu J, Huo S, Kumar A, Wei T, Zhang X, Jin S, Gan Y, Wang PC. 2012. Size-dependent localization and penetration of ultrasmall gold nanoparticles in cancer cells, multicellular spheroids, and tumors in vivo. ACS Nano, 6:4483-4493.
Huang Y, Hong J, Zheng S, Ding Y, Guo S, Zhang H, Zhang X, Du Q, Liang Z. 2011. Elimination pathways of systemically delivered siRNA. Molecular Therapy, 19:381-385.
Jackson AL, Burchard J, Schelter J, Chau BN, Cleary M, Lim L, Linsley PS. 2006. Widespread siRNA “off-target” transcript silencing mediated by seed region sequence complementarity. RNA, 12:1179-1187.
Jarad G, Miner JH. 2009. Update on the glomerular filtration barrier. Current Opinion in Nephrology and Hypertension, 18:226-232.
Judge AD, Sood V, Shaw JR, Fang D, McClintock K, MacLachlan I. 2005. Sequence-dependent stimulation of the mammalian innate immune response by synthetic siRNA. Nature Biotechnology, 23:457-462.
Kakizawa Y, Furukawa S, Ishii A, Kataoka K. 2006. Organic-inorganic hybrid-nanocarrier of siRNA constructingthrough the self-assembly of calcium phosphate and PEG-based block aniomer. Journal of Controlled Release, 111:368-370.
Kessenbrock K, Plaks V, Werb Z. 2010. Matrix metalloproteinases: Regulators of the yumor microenvironment. Cell, 141:52-67.
Kim T, Hyeon T. 2014. Applications of inorganic nanoparticles as therapeutic agents. Nanotechnology, 01:2001.
Kobayashi H, Watanabe R, Choyke PL. 2014. Improving conventional enhanced permeability and retention (EPR) effects; What is the appropriate target? Theranostics, 4:81-89.
Konishi M, Kawamoto K, Izumikawa M, Kuriyama H, Yamashita T. 2008. Gene transfer into guinea pig cochlea using adeno-associated virus vectors. Journal of Gene Medicine, 10:610-618.
Li J, Xue S, Mao ZW. 2016. Nanoparticle delivery systems for siRNA-based therapeutics. J. Mater. Chem, 4:6620-6639.
Li J, Yang Y, Huang L. 2012. Calcium phosphate nanoparticles with an asymmetric lipid bilayer coating forsiRNA delivery to the tumor. Journal of Controlled Release, 158:108–114.
Li J, Yu X, Wang Y, Yuan Y, Xiao H, Cheng D, Shuai X. 2014. A reduction and pH dual-sensitive polymeric vector for long-circulating and tumor-targeted siRNA delivery. Advanced Materials, 26:8217–8224.
Li L, Wang R, Wilcox D, Zhao X, Song J, Lin X, Kohlbrenner WM, Fesik SW, Shen Y. 2012. Tumor vasculature is a key determinant for the efficiency of nanoparticle-mediated siRNA delivery. Gene therapy, 19:775-780.
Li W, Hu ZF, Chen B, Ni GX. 2013. Response of C2C12 myoblasts to hypoxia: The relative roles of glucose and oxygen in adaptive cellular metabolism. BioMed Research International, 326-346.
Martinez J, Patkaniowska A, Urlaub H, Lührmann R, Tuschl T. 2002. Single-stranded antisense siRNAs guidetarget RNA cleavage in RNAi. Cell, 110:563-574.
Meade BR, Dowdy SF. 2007. Exogenous siRNA delivery using peptide transduction domains/cell penetrating peptides. Advanced Drug Delivery Reviews, 59:134-140.
Meister G, Tuschi T. 2004. Mechanisms of gene silencing by double-stranded RNA. Nature, 431:343-349.
Mindell JA. 2012. Lysosomal acidification mechanisms. Annual Review of Physiology, 74:69-86.
Moghimi SM, Hunter AC, Murray JC. 2001. Long-circulating and target-specific nanoparticles: Theory to practice. Pharmacological Reviews, 53:283-318.
Pai S, Lin YY, Macaes B, Meneshian A, Hung CF, Wu TC. 2006. Prospects of RNA interference therapyfor cancer. Gene Ther, 13:464-477.
Pittella F, Zhang M, Lee Y, Kim HJ, Tockary T, Osada K, Ishii T, Miyata K, Nishiyama N, Kataoka K. 2011. Enhanced endosomal escape of siRNA-incorporating hybrid nanoparticles from calciumphosphate and PEG-block charge-conversional polymer for efficient gene knockdown with negligiblecytotoxicity. Biomaterials, 32:3106-3114.
Qiu S, Adema CM, Lane T. 2005. A computational study of off-target effects of RNA interference. Nucleic Acids Res, 33:1834-1847.
Resnier P, Montier T, Mathieu V, Benoit JP, Passirani C. 2013. A review of the current status of siRNAnanomedicines in the treatment of cancer. Biomaterials, 34:6429-6443.
Rozema DB, Lewis DL, Wakefield DH, Wong SC, Klein JJ, Roesch PL, Bertin SL, Reppen TW, Chu Q, Blokhin AV. 2007. Dynamic PolyConjugates for targeted in vivo delivery of siRNA to hepatocytes. Proc. Natl. Acad. Sci. USA, 104:12982-12987.
Sashital DG, Doudna JA. 2010. Structural insights into RNA interference. Curr.Opin.Struct. Biol, 20:90-97.
Sonawane ND, Szoka FC, Verkman AS. 2003. Chloride accumulation and swelling in endosomes enhances DNA transfer by polyamine-DNA polyplexes. Free Radical Biology & Medicine, 278:44826-44831.
Van de Veire S, Stalmans I, Heindryckx F, Oura,H, Tijeras-Raballand A, Schmidt T, Loges S, Albrecht I, Jonckx B, Vinckier S. 2010. Pharmacological and genetic evidence for the efficacy of PlGFinhibition in cancer and eye disease. Cell, 141:178-190.
Vaupel P, Kallinowski F, Okunieff P. 1989. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res, 49:6449-6465.
Wang J, Mi P, Lin G, Wáng YXJ, Liu G, Chen X. 2015. Imaging-guided delivery of RNAi for anticancertreatment. Advanced Drug Delivery Reviews, 104:44-60.
Wartiovaara J, Öfverstedt LG, Khoshnoodi J, Zhang J, Mäkelä E, Sandin S, Ruotsalainen V, Cheng RH, Jalanko H, Skoglund U. 2004. Nephrin strands contribute to a porous slit diaphragm scaffold as revealed by electron tomography. J. Clin. Investig, 114:1475-1483.
Wu SY, Lopez-Berestein G, Calin GA, Sood AK. 2014. RNAi Therapies: Drugging the Undruggable. Science Translational Medicine, 6:240-247.
Young SWS, Stenzel M, Yang JL. 2016. Nanoparticle-siRNA: A potential cancer therapy? Crit. Rev. Oncol. Hematol, 98:159-169.
Zhang L, Li Y, Yu JC. 2014. Chemical modification of inorganic nanostructures for targeted and controlled drug delivery in cancer treatment. J. Mater. Chem. B, 2:452-470.
Zhao M, Sun J, Zhao Z. 2013. Synergetic regulatory networks mediated by oncogene-driven microRNAs andtranscription factors in serous ovarian cancer. Molecular BioSystems, 9:3187-3198.





